
만일, 다수의 연속적이고도 경미한 변화에 의해서는 결코 형성될 수 없는 그 어떤 복잡한 기관이라도 이 세상에 존재한다는 게 증명된다면,
나의 학설은 붕괴할 것이다.
나는 아직 그런 사례를 발견하지 못했다.
― 찰스 다윈, 종의 기원
“음, 이제 모듈(LEM)에서 내려가겠다.”
“한 사람에게는 작은 발걸음,
인류에게는 커다란 도약이다.”
직립 이족 보행을 두고 ‘위험이 따르는 사업’이라 말했던 존 네이피어 또한, 그 사업이 단 하나의 이유만으로 진척되진 않았을 것이라 덧붙였다. 이족 보행은 엄청나게 극적이면서도 수많은 인과가 한데 얽힌 대사업이었고, 또한 수많은 단점과 장점이 한 데 뒤엉켜있던 인류 최초의 대실험이나 다름없었다.
이 실험이 성공적으로 끝났다는 조건 하에서야 비로소 최초의 ‘인류’가 출현할 수 있었으니까.

그리고 두 발로 걷는 모양새가 좋든 싫든, 우리 모두는 그 실험 결과물이며, 마침내 밤하늘에 떠있던 달 위에까지 발걸음을 남기기에 이르렀다. 오늘은 우리가 어째서, 그리고 어떻게 두 발로 걷기 시작했는지 이야기할 것이다.
두 발로 걷기 시작한 초기 호미닌
지난 시간에 우리는 사헬란트로푸스 차덴시스, 오로린 투게넨시스가 최초의 호미닌 후보이며, 최초의 이족보행자인 동시에 생의 대부분을 나무에서 보냈다는 사실을 살펴보았다. 이들은 오늘날과는 다른 풍경의 동부아프리카에 살았다.


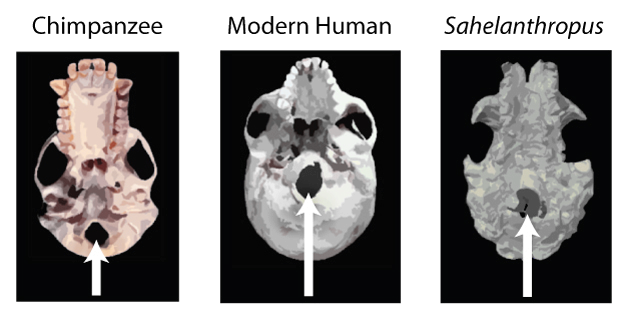
700~600만 년 전의 이들은 여러모로 인간적이지 못했다. 뾰족하고 날카로운 송곳니를 가지고 있었고, 성적 이형성은 컸으며, 과일과 나뭇잎을 주로 먹는 나무 위의 일상을 보냈으니까. 하지만 이들이 호미닌이 된 이유는 따로 있다. 이들의 대퇴골(Femur)과 대후두공(Foramen magnum)의 구조가 우리들과 서서히 닮아가고 있었기 때문이다. 그리고 그것은, 이들이 걷기 시작했다는 사실을 암시한다.
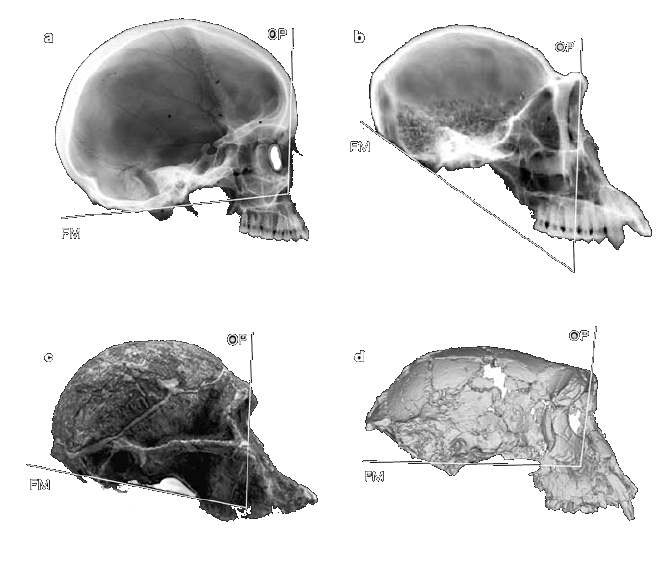
a 호모 사피엔스(우리)
b 침팬지
c 오스트랄로피테쿠스 아프리카누스
d TM 266(사헬란트로푸스 차덴시스)
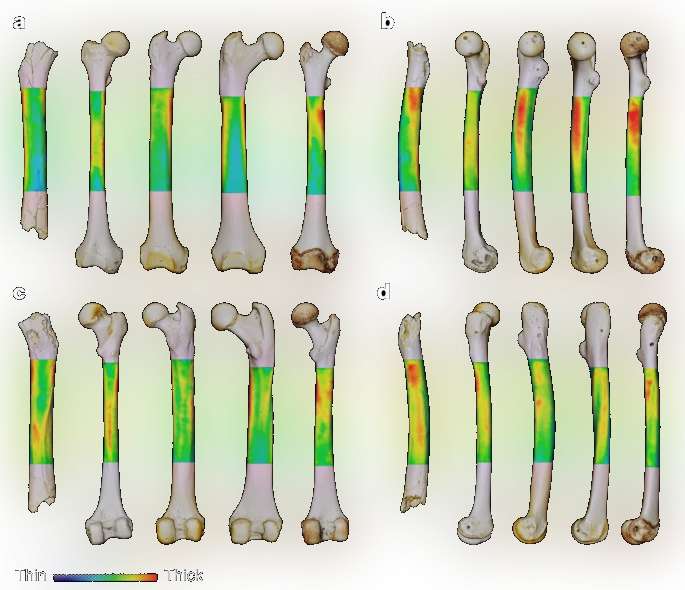
그들의 대퇴골을 현존하는 유인원류와 비교할때, 특히 후면(c)과 측면(d) 부분은 우리, 즉 현생인류와 가장 흡사하며, 대퇴골간(shaft of femur)의 기하학적 특징 또한 이들이 능히 두 발로 걷는 데 적응했다는 사실을 암시한다. 사헬란트로푸스 차덴시스, 오로린 투게넨시스에게서 공통적으로 보이는 직립 보행과 관련된 특징들은 적어도 그들이 걷는 데에 있어서 침팬지보다 우리와 가까웠다는 사실을 말해주는 것이다.
한마디로 이들은, 침팬지와의 공통조상으로부터 우리 쪽을 향해 뻗기 시작한 한 갈래 줄기였다.
이 별것도 아닌 것처럼 보이는 사소한 차이점이 곧 우리가 갖고 있는 대부분이며, 우리의 초기 진화를 말해주는 가장 강력한 증거다.
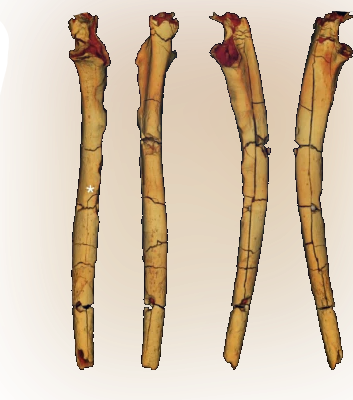
반면, 그들의 척골(Ulnar) 형태는 그들의 팔이 여전히 나무를 타기 위해 최적화 돼있었으리라는 것을 암시한다. 그들의 팔뚝과 팔꿈치는, 마치 무언가에 찰싹 달라붙어 있거나 기어 오르는데 특화돼있는 것처럼 보이며, 팔만 보면 우리보다는 차라리 오랑우탄에 가까웠다. 이를 통해 그들이 여전히 생의 대부분을 나무에서 보냈을 것이라 추측할 수 있다. 하지만 그럼에도, 그들은 이따금 두 발로 똑바로 서서 비뚝비뚝 걸었다. *

* 최신 연구결과(Roberto, Aude, Damiano, & Bernard (2020))는 TM 266(사헬란트로푸스 차덴시스) 대퇴골 형태가 이족보행의 증거로서는 불충분하며, 따라서 초기 호미닌도 아니라고 주장한다. (이렇게 정반대의 학설들이 난립하는 것은 발굴된 화석자료들의 양과 질 모두가 빈약한 데에 기인한다.) 반면, 100만 년 뒤 후배 뻘인 오로린 투게넨시스는 그들의 대퇴골 구조에서 볼 수 있는 패턴들이 이후 오스트랄로피테쿠스를 비롯한 뒷 세대의 습관적 직립이족보행(habitual bipedality) 호미닌들의 구조와 연속성을 띠고 있기 때문에, (사헬이 아니라면) 그들이 이족보행의 명백한 선조라는 데에 별다른 이견이 없다.
그런데 이들은 도대체 왜! 걷기 시작한걸까?
이족보행의 단점
두 발로 걷는다는 것은, 단순히 직립을 가능케 하는 것을 넘어서, 직립한 뒤에 ‘보행’한다는 것을 의미한다. 오늘날의 침팬지도 일시적으로는 직립보행을 할 수 있지만 그 모습은 상당히 어색하다. 침팬지는 이 때, 무릎을 구부린 채 뒤뚱뒤뚱 걷는데, 이 모습을 지금 따라해보면, 두 발로 걷는게 얼마나 힘이 드는지 단번에 이해할 수 있다. 침팬지가 걷는 것은 우리가 스쿼트 도중의 자세로 걸어다니는 것과 다를 바가 없는 것이다. 무릎 대신 근육이 균형을 지탱해야하는데, 이게 굉장히 힘들다*
*직접 따라해봤는데 뭔가 원숭이 된 기분이었습니다.
침팬지가 잠깐이나마 걸을 수 있는 것은 나무 위에서 꼬리 대신 손을 뻗어 열매를 먹기 위해 몸을 길게 늘일 필요가 있었기 때문이지, 나무 아래로 내려와 뒤뚱뒤뚱 오랫동안 걸어다니기 위함이 아니다. 그들은 땅에서 직립보행 대신 ‘너클보행’을 한다.
그들이 직립보행을 할 땐, 고개도 정면을 주시하기 힘들다. 저도 무릎 굽힌 채 앞을 보니까 목이 너무 아팟어요. 즉, 직립보행을 편안하게 하기 위해서는 단순히 골반이나 발목 발가락 뼈 몇개가 다른 수준으론 안되고, 말 그대로 머리부터 발끝까지 완전히 다른 뼈 구조를 지녀야하며 당연히 그에 맞춰 근육의 위치도 따라 변해야한다.
무엇보다, 대후두공의 위치가 두개골의 뒤편이 아니라 중심에 있어야한다.
전 대후두공이 중심에 있는 현대인인데도 무릎 굽힌 채로는 걷기 힘들더라구요,,,
아니, 그래서 단점투성인데 도대체 왜 두 발로 걷게된 거냐구...!!!
이족보행의 장점
아무튼 이렇게 불편하고 얻기도 힘든 능력에 무슨 장점이 있길래 호미닌들은 일어서게 된 걸까? 그리고 어째서 궁극적으로는 나무 아래로 완전히 내려와 오랫동안 두 발로만 걷게 된걸까? 사실 나무 위에서도 두 발로 걷는 행위 자체는 단점만큼이나 장점도 꽤 많다. 당연히... 뭐라도 장점이 있으니까 살아남았을 것이다...
1. 효율적 보행자 가설(Efficient Walker Hypothesis)

아까 전에 언급했듯, 나무 위에서 두 발로 서면, 일단 멀리있는 과일에 손을 뻗기 유리해지고, 이 숲에서 저 숲으로, 들판 아래서 나무 위로, 타잔처럼 뛰어 돌아다니기에도 훨씬 힘이 덜 들게된다. 온 몸으로 뛰어다니는 게 아니라 두 발로만 까딱까딱 돌아다니니까요. 이를 통해 숲에서 네 발로만 사는 원숭이들에 비해서 그 외 지역에서의 몇가지 먹거리 선택지가 늘어납니다.게다가, 뒤의 환경 가설에서 후술하겠지만, 이족 보행은 미어캣처럼 주위를 살피기에도 더 유리하다. 더 높은 곳에서 더 멀리까지 볼 수 있는데, 여러모로 이득이나 위협을 효과적으로 감지할 수 있었을 것입니다. 즉, 두 발로 걷는 것은 ‘숲에서도’ 유리한 부분이 있었다.

게다가, 이족 보행은 뜻밖의 이득도 가져왔습니다. 보행에 종속돼있었던 두 앞발을 두 팔로 해방시킨 일대 혁명이었다. 이제 호미닌은 해방된 두 팔로 ‘무언가’를 운반할 수 있게됐다. 어머니 호미닌은 젖먹이 새끼를 품에 안고 다님으로써, 먹이를 찾으러 나갔다온 사이를 틈 탄 포식자들이 어린아이들을 ‘실종’시킬 확률을 줄일 수 있었으며, 수컷 호미닌은 연모하는 암컷 호미닌들에게 두 손 가득 가져온 달달한 과일들을 구애의 선물로 줄 수 있었을 것이다. 그리고 먼 훗날(330만 년 전), 손에 쥔 무언가는 짱돌, 즉 도구가 되었는데, 그것에 대한 이야기는 다음에 이어서 할 것이다.
2. 러브조이의 사랑 넘치는 가설
오언 러브조이*가 1988년에 제시한 또 다른 가설은 여기에서 시작된다. 진화의 결과, 어떤 초기 호미닌들(이를테면 사헬란트로푸스 차덴시스나 오로린 투게넨시스)은 전에 비해 아주아주 약간 뇌가 커지는 동시에 나무 위에서 두 발로 더 쉽게 걸을 수 있게 골반이 변형됐다. 뇌가 커지자 자연스레 두개골도 커졌고, 골반이 비틀리자 산도는 좁아졌다. 머리가 더 커진 아이를 더 좁아진 산도로 낳으려니, 아이와 함께 죽는 산모가 늘어났고, 미숙아를 낳는 산모가 자연의 선택을 받게되었다.

이렇게 미숙한 채로 빛을 보게된 아이들은 성체가 되기전까지 외부 위협에 쉽게 노출됐을 것이다. 아기는 어미의 보살핌에 더욱 의존하게 되어, 아이를 돌보는 데 여념이 없는 어미 또한 타 개체(대부분의 경우 수컷 파트너)에게 식량을 제공받아야했을 것이다. 그런데, 대부분의 선조 영장류 짐승들이 그러했듯, 어떤 책임감없는 초기 호미닌 수컷들은 암컷이 새끼를 낳자마자(즉, 목적을 달성하자마자) 다른 암컷을 찾으러 떠나버리기를 반복했을 것이고, 그런 상황 속에서 비참하게 버려진 암컷과 새끼는 살아남기 힘들었을 것이다.
암컷들은 자연스레 새끼를 부양해줄 수 있는 다정한 수컷들, 그리고 더 많은 식량을 손쉽게 날라줄 수 있는 이족 보행에 능한 수컷들을 선호하게 됐을 것이다. 즉, 결혼(이 땐 난혼에 가까웠지만) 상대를 더 까다롭고 신중하게 골라야하는 시기가 도래했던 것이다. 이족 보행에 능한 수컷이 선택받는 과정이 반복돼, 더 많은 개체가 이족 보행에 능해질수록, 비례해 산도가 더 좁아졌을 것이고, 아이는 더욱 더 미숙한 채로 태어나게 되는 일종의 ‘되먹임고리(feedback loop)’가 형성됐다.
이 과정을 통해 점차 공격적인 수컷이 도태, 송곳니의 날카로움 정도나 암수 체격차이로 대표되는 성적 이형성이 점차 적어졌을 것이라 추측할 수 있는데, 실제로 정확히 이 시기의 사헬란트로푸스 차덴시스와 오로린 투게넨시스에게서 그런 경향이 발견된다.
*오언 러브조이는 이전(1981년)에 발표했던 가설 등, 여러모로 각종 오해와 확대해석을 불러일으킬 수 있는 자극적인 내용 등으로 비판받아왔다. 그러나 나는 러브조이 가설의 시대적 한계(나온지 40년된 가설이다)와 세부적 오류 몇가지를 인정한다 해도, 그에 대한 소위 ‘비판’ 대부분이 냉정하게 거리를 둔 정당한 비판이라기보다는 확증편향과 무분별한 혐오감정에 기반한 원색적인 ‘비난’에 가깝다고 생각하며, 그의 가설 단독으로보다는 다른 가설과 결합했을 때, 여전히 유효하다고 생각한다.

사족이지만, 여기까지만 봐도 두 발로 걷는 게 이렇게나 좋은데 왜 호미닌 이외의 다른 사족보행 동물들, 이를테면 고양잇과 맹수들이나 우제류 초식동물들은 두 발로 걷는 길을 가지 않았을까?
그건 이들이 나무를 타지 않았기 때문이다. 이족보행에 적합하도록 진화하는 길은, 하물며 나무 위에서 비뚝비뚝 오랜 ‘전적응’의 시간을 보냈던 호미닌에게도 끔찍한 시련*을 강요했다.
* 이 시련은 지금까지도 현재진행형인데, 아마 저를 비롯해 각종 디스크로 고통받고 계시는 분들은 제 말에 온몸으로 동의하실 것 같네요...

그런데 나무도 안타고 숲 대신 사바나 초원을 거닐던 사자나 사슴은, 온 몸의 척추와 근육을 모조리 바꿔 두 발로 갑자기 걸어다니는 급가속대회전드리프트보다는 (예를 들자면) 송곳니나 뿔의 크기를 키우는 쪽이 여러모로 효율적이었을 것이다. 더욱이, 본래 식물을 주식으로 삼고 남이 먹다남긴 고기 찌꺼기기를 곁들일 뿐이었던 호미닌들과 달리, 빠르게 달려 사냥감을 잡아먹어야하는 포식자와 그로부터 달아나야하는 피식자는 순간 가속에 유리한 네 발을 발달시키는 편이 단기적으로 더 유리했을 것이다.
환경 가설
대부분의 영장류는 어디 살까?


숲이다. 정확히는 밀림으로 빼곡한 열대 우림, 즉 상록 우림이다.

또 어떤 영장류, 이를테면 침팬지들은 개방 삼림지역에서도 곧잘 살아간다.

그리고, 사바나 관목지(Shrubland)에 사는 영장류도 있다.* 수백만 년 뒤(지금으로부터 200만 년 전, 그러니까 사헬이나 오로린보다 500만년이나 뒤 쯤?)의 호미닌도 훗날 이 대열에 합류할 것이다. 그리고 이족 보행의 완성또한 이 사바나로 떠난 여행과 상당한 연관이 있을 것이다. 사바나가 이족 보행을 낳지는 않았겠지만, 적어도 이족 보행은 사바나를 마음의 고향으로 택한 것이다.
*개코원숭이, 파타스원숭이 등등. 관목지는 대체로 나무 높이가 3m를 넘지않는 지역이다.
사바나 가설

지난 시간에 잠시 지나가듯이 언급했던 동아프리카 지구대 형성 사건은 히말라야 산맥이 솟고, 인도네시아의 해로가 막힌 사건과 함께 초창기 인류 진화에 큰 영향을 미쳤다.
이로인해 300만~400만년 전부터 본격적으로 비그늘 현상이 동아프리카 내륙 지역에 나타나 가물게 되었고, 이곳의 열대우림은 점차 마르기 시작해, 서서히 건조한 바위투성이 사막 관목지 배경의 사바나 기후로 변해가고 있었다. 그리고 이 사바나 기후에 적응한 동식물들이 새롭게 번성하기 시작한다.
영양과 얼룩말같은 풀 뜯어먹는 거대한 초식동물들이 초원을 메웠고, 그에 따라 이들을 잡아먹는 대형 육식동물들도 번성했다. 사바나 곳곳에 초식동물의 반쯤 썩은 사체들(사체가 생기는 원인은 포식자에 의한 피살보다 자연사가더 많았다)이 생겨났고, 또 그 틈은 청소 동물들의 새로운 기회가 되었다. 그리고 훗날 어떤 호미닌들도 이 틈에 합류, ‘사바나의 청소부’를 자처할 것이다.
이러한 변화의 과도기에, 이 부근의 기후와 자연환경, 그리고 생물학적 다양성은 매우 복잡해졌다. 숲 바로 옆에 너른 들판이 있었고, 그 바로 옆에 높디높은 설산이 있었고, 그 바로 옆에는 고원이 있었고, 그 옆에는 가파른 골짜기가 있었다. 조금만 위도와 경도를 옮겨도 전혀 다른 풍경이 펼쳐지고 해발고도가 휙휙 바뀌는 이 곳은 가혹하고도 매력적인 기회의 땅이자, 진화의 가속실험장이었다. 이중에서 특히 초기 호미닌의 주 무대로 주목되는 곳은 사바나 초원과 삼림 지역 사이에 끼인 이른바, ‘모자이크’ 지역이었다.
수많은 종이 하루아침(사실은 십수만 년 사이)에 멸종당했고, 다시 수많은 집단이 새로운 틈새시장을 발견했다. 어떤 무리들은 무식하게도 거대한 체격과 튼튼한 턱을, 또 어떤 무리들은 비리비리한 체구와 기괴하게 생긴 대두를 발달시켰다. 그리고 새로운 환경에 적응하며, 다양한 호미닌 무리가 하루아침에 옛 전통과 육신을 버리고 새롭게 탈바꿈, 즉 발산했다. 이족 보행 또한 그 탈바꿈을 통해 마침내 완성된 하나의 테크트리 세트였다.
이족보행은 뜨거운 사바나의 열기를 더 좁은 면적으로 받게 해주었고, 무엇보다 신체의 윗부분을 지면으로부터 일렁이는 지열에서 멀어질 수 있도록 했으며, 대평야를 지배하는 무시무시한 신체스펙의 포식자들을 더 잘 발견할 수 있도록 도왔다.
하지만 동부아프리카가 사바나로 변해버리는 건 먼 훗날의 이야기다. 환경은 하루아침에 변하지 않았고, 숲을 따라 피난하는 것도 그들 스스로 전혀 의식하지 못할 정도로 느릿느릿 벌어졌을 것이다. 아직 어색하게 이족보행하는 호미닌들에게, 굳이 위험으로 가득한 사바나를 어슬렁거리는 것보다 수분기 있는 옆동네 땅과 숲에서 살아갈 기회는 여전히 많았다. 아직, 인간은 사바나가 아니라 숲에 살았다. 우리가 살펴보고 있는 이 시기(700~600만년 전)의 사바나는 여전히 이따금 두 발로 잘 걷는 어떤 용감한 호미닌 집단이 도전했다가, 잡아먹혀버리고 마는 호미닌의 무덤에 가까웠으리라.
결론
시계를 거꾸로 돌리면 돌릴수록, 우리의 조상들은 그 어떤 ‘인간적’ 특성도 거저 얻지는 못했다는 것을, 그리고 그 대부분의 특성들이 다른 수많은 경쟁자들과 함께, 아주 작게 꾸물거리고만 있었다는 것을 알 수 있다. 오늘날의 우리가 인간성과 결부돼있다고 여기는 대표적 특성들은 700만년전엔 생태적 압력이라는 벽에 간신히 붙어 헐떡이는 존재들이었다. 어떤 특성이 얼마나 더 빠르게, 그리고 어떤 방향으로 바뀔지는 다만 외부의 환경과 호미닌 내부에 도사린 발생학적 특징이 어떻게 상호작용하는지에 달려 있을 뿐이었다.
나는 물가의 숲에 살았던 초기 호미닌들 중 적어도 한 갈래가, 위에서 언급한 (그리고 위에서 언급하지 않았지만 수없이 많은 그럴듯한) 가설들 사이의 연쇄작용에 자연스레 이끌려 이족 보행을 더 정교하게 발달시키는 방향으로 나아가게 되었을 것이라 생각한다. 그리고 그들은 수백만년 간 네 발로 걷는 친척들과 부대끼며 마침내 우리에까지 이르렀을 것이다.
요약:
가장 먼저, 900~700만년 전 사이, 어떤 숲의 어떤 무리들은 숲을 두 발로 이동하는 것의 이점을 살리기 시작했다.
그때부터 그들은 훗날 침팬지가 되는 부류의 조상과 조금씩 갈라졌고,
한 번 똑바로 서서 다른 길을 걷기 시작한 호미닌들은 자연스레 형성된 여러 되먹임고리들에 의해 이족 보행으로 향하는 일방통행이 강제되었으며,
300~200만 년 전 쯤 멋모르고 들어서버린 사바나 풍 초행길로 인해,
수백만 년이 지난 오늘날까지 (두 발로) 걷게 된 것이다.
다윈의 말마따나 두 발로 걷는 것은 두 손을 해방시켰고,
무엇보다 궁극적으로는, 숲의 사람들이 초원의 떠돌이가 될 수 있도록 도왔다.
나무 위의 생활은 이족보행의, 이족보행은 도구사용의, 그리고 도구 사용은 지능 발달의, 전적응이 되었다.
이족보행을 통해 우리는 진정한 의미의 '인류'사, 그 첫 단추를 끼웠다.
다음 시간엔, 나무 아래서 두 발로 어색하게 걷기 시작한 초기 호미닌이 어떻게 점차 새로운 사바나 환경에 적응해나가는지를 알아볼 것이다.
참고:
Zollikofer, C. P. E. et al. Virtual cranial reconstruction of Sahelanthropus tchadensis. Nature 434, 755-759 (2005)
Guy, F. et al. Morphological affinities of the Sahelanthropus tchadensis (late Miocene hominin from Chad) cranium. Proceedings of the National Academy of Sciences of the United States of America 102, 18836-18841 (2005)
University of Arizona. "Study Identifies Energy Efficiency As Reason For Evolution Of Upright Walking." ScienceDaily. ScienceDaily, 17 July (2007)
Wood, B., Harrison, T. The evolutionary context of the first hominins. Nature 470, 347–352 (2011)
M. Bleuze, Proximal femoral diaphyseal cross-sectional geometry in Orrorin tugenensis, HOMO, Volume 63, Issue 3, (2012)
Almécija, S., Tallman, M., Alba, D. et al. The femur of Orrorin tugenensis exhibits morphometric affinities with both Miocene apes and later hominins. Nat Commun 4, 2888 (2013)
Hannah M. Liddy, Sarah J. Feakins, Jessica E. Tierney. Cooling and drying in northeast Africa across the Pliocene. Earth and Planetary Science Letters. Volume 449. (2016)
Roberto Macchiarelli, Aude Bergeret-Medina, Damiano Marchi, Bernard Wood, Nature and relationships of Sahelanthropus tchadensis, Journal of Human Evolution, Volume 149 (2020)
Daver, G., Guy, F., Mackaye, H.T. et al. Postcranial evidence of late Miocene hominin bipedalism in Chad. Nature 609, 94–100 (2022)
Zeller, Elke et al. “Human adaptation to diverse biomes over the past 3 million years.” Science (New York, N.Y.) vol. 380,6645 (2023)
Peter A. Stamos, Zeresenay Alemseged, Hominin locomotion and evolution in the Late Miocene to Late Pliocene, Journal of Human Evolution, Volume 178 (2023)
'내글 > 역사' 카테고리의 다른 글
| 삼국지 황건적 두목 '장각'은 한낱 사이비 교주였을까 (0) | 2024.06.27 |
|---|---|
| 삼국지 황건의 난이 로마 제국 탓인 이유 (0) | 2024.06.27 |
| "’인면수심’의 조상님..." , 후손들은 결국 ‘파묘’했다. (0) | 2024.04.02 |
| "조상님이 사실 햄스터?" 최초의 인류, 사실 찍찍거렸다? (0) | 2024.03.30 |
| 중세 수레바퀴 고문 (6) | 2023.07.18 |